Еще в 20-х гг. А. Лотка (Lotka), а несколько позднее независимо от него В. Вольтерра предложили математические модели, описывающие сопряженные колебания численности популяций хищника и жертвы. Рассмотрим самый простой вариант модели Лотки-Вольтерра. В основе модели лежит ряд допущений:
1) популяция жертв в отсутствие хищника растет экспоненциально,
2) пресс хищников тормозит этот рост,
3) смертность жертв пропорциональна частоте встреч хищника и жертвы (или иначе, пропорциональна произведению плотностей их популяций);
4) рождаемость хищника зависит от интенсивности потребления жертв.
Мгновенная скорость изменения численности популяции жертв может быть выражена уравнением
dN ж /dt = r 1 N ж - p 1 N ж N х,
где r 1 - удельная мгновенная скорость популяционного роста жертвы, p 1 - константа, связывающая смертность жертв с плотностью хищника, a N ж и N х - плотности соответственно жертвы и хищника.
Мгновенная скорость роста популяции хищника в этой модели принимается равной разности рождаемости и постоянной смертности:
dN х /dt = p 2 N ж N х – d 2 N х,
где p 2 - константа, связывающая рождаемость в популяции хищника с плотностью жертв, a d 2 - удельная смертность хищника.
Согласно приведенным уравнениям каждая из взаимодействующих популяций в своем увеличении ограничена только другой популяцией, т.е. рост числа жертв лимитируется прессом хищников, а рост числа хищников - недостаточным количеством жертв. Никакого самоограничения популяций не предполагается. Считается, например, что пищи для жертвы всегда достаточно. Также не предполагается и выхода популяции жертв из-под контроля хищника, хотя на самом деле такое бывает достаточно часто.
Несмотря на всю условность модели Лотки-Вольтерра, она заслуживает внимания уже хотя бы потому, что показывает, как даже такая идеализированная система взаимодействия двух популяций может порождать достаточно сложную динамику их численности. Решение системы этих уравнений позволяет сформулировать условия поддержания постоянной (равновесной) численности каждого из видoв. Популяция жертв сохраняет постоянную численность, если плотность хищника равна r 1 /p 1, а для того чтобы постоянство сохраняла популяция хищника, плотность жертв должна быть равна d 2 /p 2 . Если на графике отложить по оси абсцисс плотность жертв N ж , а по оси ординат - плотность хищника N х , то изоклины, показывающие условие постоянства хищника и жертвы, будут представлять собой две прямые, перпендикулярные друг другу и координатным осям (рис. 6,а). При этом предполагается, что ниже определенной (равной d 2 /p 2) плотности жертв плотность хищника всегда будет уменьшаться, а выше - всегда увеличиваться. Соответственно и плотность жертвы возрастает, если плотность хищника ниже значения, равного r 1 /p 1 , и уменьшается, если, она выше этого значения. Точка пересечения изоклин соответствует условию постоянства численности хищника и жертвы, а другие точки на плоскости этого графика совершают движение по замкнутым траекториям, отражая, таким образом, регулярные колебания численности хищника и жертвы (рис. 6,б). Размах колебаний определяется начальным соотношением плотностей хищника и жертвы. Чем ближе оно к точке пересечения изоклин, тем меньше окружность, описываемая векторами, и соответственно меньше амплитуда колебаний.
Рис. 6. Графическое выражение модели Лотки-Вольтера для системы хищник-жертва.
Одна из первых попыток получения колебаний численности хищника и жертвы в лабораторных экспериментах принадлежала Г.Ф. Гаузе (Gause). Объектами этих экспериментов были инфузория парамеция (Paramecium caudatum ) и хищная инфузория дидиниум (Didinium nasutum ). Пищей для парамеции служила регулярно вносимая в среду взвесь бактерий, а дидиниум питался только парамециями. Данная система оказалась крайне неустойчивой: пресс хищника по мере увеличения его численности приводил к полному истреблению жертв, после чего вымирала и популяция самого хищника. Усложняя опыты, Гаузе устраивал убежище для жертвы, внося в пробирки с инфузориями немного стеклянной ваты. Среди нитей ваты могли свободно перемещаться парамеции, но не могли дидиниумы. В таком варианте опыта дидиниум съедал всех парамеций, плавающих в свободной от ваты части пробирки, и вымирал, а популяция парамеции затем восстанавливалась за счет размножения особей, уцелевших в убежище. Некоторого подобия колебаний численности хищника и жертвы Гаузе удалось добиться только в том случае, когда он время от времени вносил в культуру и жертву и хищника, имитируя таким образом иммиграцию.
Через 40 лет после работы Гаузе его опыты были повторены Л. Лакинбиялом (Luckinbill), использовавшим в качестве жертвы инфузорию Paramecium aurelia , а в качестве хищника того же Didinium nasutum . Лакинбиллу удалось получить несколько циклов колебаний численности этих популяций, но только в том случае, когда плотность парамеций была лимитирована нехваткой пищи (бактерий), а в культуральную жидкость добавляли метилцеллюлозу - вещество, снижающее скорость движения как хищника, так и жертвы и потому уменьшающее частоту их возможных встреч. Оказалось также, что добиться колебаний хищника и жертвы легче, если увеличить объем экспериментального сосуда, хотя условие пищевого лимитирования жертвы и в этом случае обязательно. Если же к системе сосуществующих в колебательном режиме хищника и жертвы добавляли избыточную пищу, то ответом был, быстрый рост численности жертвы, за которым следовало возрастание численности хищника, приводящее в свою очередь к полному истреблению популяции жертвы.
Модели Лотки и Вольтерры послужили толчком для разработки ряда других более реалистичных моделей системы хищник-жертва. В частности, довольно простая графическая модель, анализирующая соотношение разных изоклин жертвы хищника, была предложена М. Розенцвейгом и Р. Мак-Артуров (Rosenzweig, MacArthur). Согласно этим авторам, стационарная (= постоянная) численность жертвы в координатных осях плотности хищника и жертвы может быть представлена в виде выпуклой изоклины (рис. 7,а). Одна точка пересечения изоклины сосью плотности жертвы соответствует минимальной допустимой плотности жертвы (нижеее популяция подвержена очень большому риску вымирания хотя бы из-за малой частоты встреч самцов и самок), а другая - максимальной, определяемой количеством имеющейся пищи или поведенческими особенностями самой жертвы. Подчеркнем, что речь идет пока о минимальной и максимальной плотностях в отсутствие хищника. При появлении хищника и увеличении его численности минимальная допустимая плотность жертвы, очевидно, должна быть выше, а максимальная - ниже. Каждому значению плотности жертвы должна соответствовать некоторая плотность хищника, при которой достигается постоянство популяции жертвы. Геометрическое место таких точек и есть изоклина жертвы в координатах плотности хищника и жертвы. Векторы, показывающие направление изменения плотности жертвы (ориентированные горизонтально), имеют разную направленность по разные стороны от изоклины (рис. 7,а).

Рис. 7. Изоклины стационарных популяций жертвы (а) и хищника (б).
Для хищника в тех же координатах также построена изоклина, отвечающая стационарному состоянию его популяции. Векторы, показывающие направление изменения численности хищника, ориентированы вверх или вниз в зависимости от того, по какую сторону от изоклины они находятся. Форма изоклины хищника, показанная на рис. 7,б. определяется, во-первых, наличием некоторой минимальной плотности жертвы, достаточной для поддержания популяции хищника (при более низкой плотности жертвы хищник не может увеличивать свою численность), а во-вторых, наличием некоторой максимальной плотности самого хищника, при превышении которой численность будет снижаться независимо от обилия жертв.

Рис. 8. Возникновение колебательных режимов в системе хищник-жертва в зависимости от расположения изоклин хищника и жертвы.
При совмещении изоклин жертвы и хищника на одном, графике возможны три различных варианта (рис. 8). Если изоклина хищника пересекает изоклину жертвы в том месте, где она уже снижается (при высокой плотности жертв), векторы, показывающие изменение численности хищника и жертвы, образуют траекторию, закручивающуюся во внутрь, что соответствует затухающим колебаниям численности жертвы и хищника (рис. 8,а). В том случае, когда изоклина хищника пересекает изоклину жертвы в ее восходящей части (т.е. в области низких значений плотности жертв), векторы образуют раскручивающуюся траекторию, а колебания численности хищника и жертвы происходят соответственно с возрастающей амплитудой (рис. 8,б). Если же изоклина хищника пересекает изоклину жертвы в области ее вершины, то векторы образуют замкнутый круг, а колебания численности жертвы и хищника характеризуются стабильной амплитудой и периодом (рис. 8,в).
Иными словами, затухающие колебания соответствуют ситуации, при которой хищник ощутимо воздействует на популяцию жертв, достигнувшую только очень высокой плотности (близкой к предельной), а колебания возрастающей амплитуды возникают, тогда, когда хищник способен быстро увеличивать свою численность даже при невысокой плотности жертв и таким образом быстро ее уничтожить. В других вариантах своей модели Poзенцвейг и Мак-Артур показали, что стабилизировать колебания хищник-жертва можно, введя «убежище», т.е. предположив, что в области низкой плотности жертв существует область, где численность жертвы растет независимо от количества имеющихся хищников.
Стремление сделать модели более реалистичными путем их усложнения проявилось в работах не только теоретиков, но и экспериментаторов. В частности, интересные результаты были получены Хаффейкером (Huffaker), показавшим возможность сосуществования хищника и жертвы в колебательном режиме на примере мелкого растительноядного клеща Eotetranychus sexmaculatus и нападающего на него хищного клеща Typhlodromus occidentalis . В качестве пищи для растительноядного клеща использовали апельсины, помещенные на подносы с лунками (вроде тех, что используются для хранения и перевозки яиц). В первоначальном варианте на одном подносе было 40 лунок, причем в некоторых из них находились апельсины (частично очищенные от кожуры), а в других - резиновые мячики. Оба вида клещей размножаются партеногенетически очень быстро, и поэтому характер их популяционной динамики можно выявить за сравнительно короткий срок. Поместив на поднос 20 самок растительноядного клеща Хаффейкер наблюдал быстрый рост его популяции, которая стабилизировалась на уровне 5-8 тыс. особей (в расчете на один апельсин). Если к растущей популяции жертвы добавляли несколько особей хищника, то популяция последнего быстро увеличивала свою численность и вымирала, когда все жертвы оказывались съеденными.
Увеличив размер подноса до 120 лунок, в которых отдельные апельсины были случайно разбросаны среди множества резиновых мячиков, Хаффейкеру удалось продлить сосуществование хищника и жертвы. Важную роль во взаимодействии хищника и жертвы, как выяснилось, играет соотношение скоростей их расселения. Хаффейкер предположил, что, облегчив передвижение жертвы и затруднив передвижение хищника, можно увеличить время их сосуществования. Для этого на подносе из 120 лунок среди резиновых мячиков располагали случайным образом 6 апельсинов, причем вокруг лунок с апельсинами были устроены преграды из вазелина, препятствовавшие расселению хищника, а для облегчения расселения жертвы на подносе были укреплены деревянные колышки, служившие своего рода «взлетными площадками» для растительноядных клещей (дело в том, что этот вид выпускает тонкие нити и с помощью их может парить в воздухе, распространяясь по ветру). В таком усложненном местообитании хищник и жертва сосуществовали в течение 8 месяцев, продемонстрировав три полных цикла колебаний численности. Наиболее важные условия этого сосуществования следующие: гетерогенность местообитания (в смысле наличия в ней пригодных и непригодных для обитания жертвы участков), а также возможность миграции жертвы и хищника (с сохранением некоторого преимущества жертвы в скорости этого процесса). Иными словами, хищник может полностью истребить то или иное локальное скопление жертв, но часть особей жертвы успеет мигрировать и дать начало другим локальным скоплениям. До новых локальных скоплений хищник рано или поздно тоже доберется, но тем временем жертва успеет расселиться в другие места (в том числе и в те, где она обитала раньше, но потом была истреблена).
Нечто подобное тому, что наблюдал Хаффейкер в эксперименте, встречается и в природных условиях. Так, например, бабочка кактусовая огневка (Cactoblastis cactorum ), завезенная в Австралию, значительно снизила численность кактуса опунции, но не уничтожила его полностью именно потому, что кактус успевает расселиться немного быстрее. В тех местах, где опунция истребляется полностью, перестает встречаться и огневка. Поэтому, когда через некоторое время сюда вновь проникает опунция, то в течение определенного периода она может произрастать без риска быть уничтоженной огневкой. Со временем, однако, огневка снова здесь появляется и, быстро размножаясь, уничтожает опунцию.
Говоря о колебаниях хищник-жертва, нельзя не упомянуть и о циклических изменениях численности зайца и рыси в Канаде, прослеженных по материалам статистики заготовок пушнины компанией Гудзон-Бэй с конца XVIII вплоть до начала XX в. Этот пример нередко рассматривался как классическая иллюстрация колебаний хищник-жертва, хотя на самом деле мы видим только следование роста численности популяции хищника (рыси) за ростом численности жертвы (зайца). Что же касается снижения численности зайцев после каждого подъема, то оно не могло объясняться только возросшим прессом хищников, а было связано с другими факторами, по-видимому, прежде всего нехваткой корма в зимний период. К такому выводу пришел, в частности, М. Джилпин (Gilpin), пытавшийся проверить, могут ли быть описаны эти данные классической моделью Лотки-Вольтерры. Результаты проверки показали, что удовлетворительного соответствия модели нет, но как ни странно, оно становилось лучше, если хищника и жертву меняли местами, т.е. трактовали рысь как «жертву», а зайца - как «хищника». Подобная ситуация нашла свое отражение и в шутливом названии статьи («Едят ли зайцы рысей?»), по сути своей очень серьезной и опубликованной в серьезном научном журнале.
Часто представители одного вида (популяции) питаются представителями другого вида.
Модель Лотки – Вольтерры – модель взаимного существования двух популяций типа «хищник – жертва».
Впервые модель «хищник – жертва» была получена А. Лоткой в 1925 году, который использовал ее для описания динамики взаимодействующих биологических популяций. В 1926 году независимо от Лотки аналогичные (к тому же более сложные) модели были разработаны итальянским математиком В. Вольтерра, глубокие исследования которого в области экологических проблем заложили фундамент математической теории биологических сообществ или т. н. математической экологии.
В математической форме предложенная система уравнений имеет вид:
где x – количество жертв, y – количество хищников, t – время, α, β, γ, δ – коэффициенты, которые отражают взаимодействия между популяциями.
Постановка задачи
Рассмотрим закрытое пространство, в котором существуют две популяции – травоядные («жертвы») и хищники. Считается, что животных не ввозят и не вывозят и что еды для травоядных животных достаточно. Тогда уравнение изменения числа жертв (только жертв) примет вид:
где $α$ – коэффициент рождаемости жертв,
$x$– размер популяции жертв,
$\frac{dx}{dt}$ – скорость прироста популяции жертв.
Когда хищники не охотятся, они могут вымирать, значит, уравнение для количества хищников (только хищников) примет вид:
Где $γ$ – коэффициент убыли хищников,
$y$ – размер популяции хищников,
$\frac{dy}{dt}$ – скорость прироста популяции хищников.
При встрече хищников и жертв (частота встреч прямо пропорциональна произведению) хищники уничтожают жертв с коэффициентом, сытые хищники могут воспроизводить потомство с коэффициентом. Таким образом, система уравнений модели примет вид:
Решение задачи
Построим математическую модель совместного существования двух биологических популяций типа «хищник – жертва».
Пусть две биологические популяции совместно обитают в изолированной среде. Среда является стационарной и обеспечивает в неограниченном количестве всем необходимым для жизни один из видов – жертвы. Другой вид – хищник – также обитает в стационарных условиях, но питается только жертвами. В роли хищников могут выступать коты, волки, щуки, лисы, а в роли жертв – куры, зайцы, караси, мыши соответственно.
Для определенности рассмотрим в роли хищников – котов, а в роли жертв – кур.
Итак, куры и коты живут в некотором изолированном пространстве – хозяйственном дворе. Среда предоставляет курам питание в неограниченном количестве, а коты питаются только курами. Обозначим через
$х$ – количество кур,
$у$ – количество котов.
Со временем количество кур и котов меняется, но будем считать $х$ и $у$ непрерывными функциями от времени t. Назовем пару чисел $х, у)$ состоянием модели.
Найдем каким образом изменяется состояние модели $(х, у).$
Рассмотрим $\frac{dx}{dt}$ – скорость изменения количества кур.
Если котов нет, то количество кур возрастает и тем быстрее, чем больше кур. Будем считать зависимость линейной:
$\frac{dx}{dt} a_1 x$,
$a_1$ – коэффициент, который зависит только от условий жизни кур, их естественной смертности и рождаемости.
$\frac{dy}{dt}$ – скорость изменения количества котов (если нет кур), зависит от количества котов y.
Если кур нет, то количество котов уменьшается (у них нет пищи) и они вымирают. Будем считать зависимость линейной:
$\frac{dy}{dt} - a_2 y$.
В экосистеме скорость изменения количества каждого вида также будем считать пропорциональным его количеству, но только с коэффициентом, зависящим от количества особей другого вида. Так, для кур этот коэффициент уменьшается с увеличением количества котов, а для котов возрастает с увеличением количества кур. Будем считать зависимость также линейной. Тогда получим систему дифференциальных уравнений:
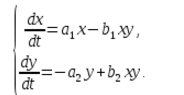
Данная система уравнений называется моделью Вольтерра-Лотки.
a1, a2, b1, b2 – числовые коэффициенты, которые называют параметрами модели.
Как видно, характер изменения состояния модели (x, y) определяется значениями параметров. Изменяя данные параметры и решая систему уравнений модели, можно исследовать закономерности изменения состояния экологической системы.
С помощью программы MATLAB система уравнений Лотки-Вольтерра решается следующим образом:
На рис. 1 представлено решение системы. В зависимости от начальных условий решения разные, чему отвечают разные цвета траекторий.
На рис. 2 представлены те же решения, но с учетом оси времени t (т.е. наблюдается зависимость от времени).
Математическое моделирование биологических процессов началось с создания первых простейших моделей экологической системы.
Допустим, в некотором замкнутом районе живут рыси и зайцы. Рыси питаются только зайцами, а зайцы – растительной пищей, имеющейся в неограниченном количестве. Необходимо найти макроскопические характеристики, описывающие популяции. Такими характеристиками являются число особей в популяциях.
Простейшая модель взаимоотношений популяций хищника и жертвы, основанная на логистическом уравнении роста, названа (как и модель межвидовой конкуренции) по имени ее создателей - Лотки и Вольтерра. Эта модель крайне упрощает исследуемую ситуацию, но все же полезна в качестве отправной точки в анализе системы хищник-жертва.
Предположим, что (1) популяция жертвы существует в идеальной (независимой от плотности) среде, где ее рост может ограничивать только наличие хищника, (2) столь же идеальна среда, в которой существует хищник, рост популяции которого ограничивает лишь обилие жертв, (3) обе популяции размножаются непрерывно согласно экспоненциальному уравнению роста, (4) скорость поедания жертв хищниками пропорциональна частоте встреч между ними, которая, в свою очередь, является функцией плотности популяций. Эти допущения и лежат в основе модели Лотки - Вольтерра.
Пусть в отсутствие хищников популяция жертвы растет экспоненциально:
dN/dt =r 1 N 1
где N -численность, а r, - удельная мгновенная скорость роста популяции жертвы. Если же хищники присутствуют, то они уничтожают особей жертвы со скоростью, которая определяется, во-первых, частотой встреч хищников и жертв, возрастающей по мере увеличения их численностей, и, во-вторых, эффективностью, с которой хищник обнаруживает и ловит свою жертву при встрече. Число жертв, встреченных и съеденных одним хищником N с, пропорционально эффективности охоты, которую мы выразим через коэффициент С 1; численности (плотности) жертвы N и времени, затраченному на поиски Т:
N C =C 1 NT (1)
Из этого выражения легко определить удельную скорость потребления жертв хищником (т.е. число жертв, поедаемых одной особью хищника в единицу времени), которую часто называют также функциональным ответом хищника на плотность популяции жертвы:
В рассматриваемой модели С 1 является константой. Это означает, что число жертв, изъятых хищниками из популяции, линейно возрастает с увеличением ее плотности (так называемый функциональный ответ типа 1). Ясно, что общая скорость поедания жертв всеми особями хищника составит:
![]() (3)
(3)
где Р - численность популяции хищника. Теперь мы можем записать уравнение роста популяции жертвы следующим образом:
При отсутствии жертвы особи хищника голодают и гибнут. Предположим также, что в этом случае численность популяции хищника будет уменьшаться экспоненциально согласно уравнению:
![]() (5)
(5)
где r 2 - удельная мгновенная смертность в популяции хищника.
Если жертвы присутствуют, то те особи хищника, которые смогут их найти и съесть, будут размножаться. Рождаемость в популяции хищника в данной модели зависит только от двух обстоятельств: скорости потребления жертв хищником и эффективности, с которой поглощенная пища перерабатывается хищником в его потомство. Если мы выразим эту эффективность через коэффициент s, то рождаемость составит:
![]()
Поскольку С 1 и s - константы, их произведение - это также константа, которую мы обозначим как С 2 . Тогда скорость роста популяции хищника будет определяться балансом рождаемости и смертности в соответствии с уравнением:
![]() (6)
(6)
Уравнения 4 и 6 вместе образуют модель Лотки-Вольтерра.
Свойства этой модели мы можем исследовать точно так же, как и в случае конкуренции, т.е. построив фазовую диаграмму, на которой численность жертвы отложена по оси ординат, а хищника - по оси абсцисс, и проведя на ней изоклины-линии, соответствующие постоянной численности популяций. С помощью таких изоклин определяют поведение взаимодействующих популяций хищника и жертвы.
Для популяции жертвы: при откуда
Таким образом, поскольку r, и С 1 , - константы, изоклиной для жертвы будет линия, на которой численность хищника (Р) является постоянной, т.е. параллельная оси абсцисс и пересекающая ось ординат в точке Р =r 1 / С 1 . Выше этой линии численность жертвы будет уменьшаться, а ниже- возрастать.
Для популяции хищника:
при откуда
Поскольку r 2 и С 2 - константы, изоклиной для хищника будет линия, на которой численность жертвы (N) является постоянной, т.е. перпендикулярная оси ординат и пересекающая ось абсцисс в точке N = r 2 /С 2 . Слева от нее численность хищника будет уменьшаться, а справа - возрастать.
Если мы рассмотрим эти две изоклины вместе, то легко заметим, что взаимодействие популяций хищника и жертвы имеет циклический характер, поскольку их численности претерпевают неограниченные сопряженные колебания. Когда велико число жертв, численность хищников растет, что приводит к увеличению пресса хищничества на популяцию жертвы и тем самым к снижению ее численности. Это снижение, в свою очередь, ведет к нехватке пищи у хищников и падению их численности, которое вызывает ослабление пресса хищничества и увеличению численности жертвы, что снова приводит к росту популяции жертвы и т.д.
Для данной модели характерна так называемая "нейтральная стабильность", которая означает, что популяции неограниченно долго совершают один и тот же цикл колебаний до тех пор, пока какое-либо внешнее воздействие не изменит их численность, после чего популяции совершают новый цикл колебаний с иными параметрами. Для того, чтобы циклы стали стабильными, популяции должны после внешнего воздействия стремиться вернуться к первоначальному циклу. Такие циклы, в отличие от нейтрально стабильных колебаний в модели Лотки-Вольтерра, принято называть устойчивыми предельными циклами.
Модель Лотки-Вольтерра, тем не менее, полезна тем, что позволяет продемонстрировать основную тенденцию в отношениях хищник-жертва возникновение циклических сопряженных колебаний численности их популяций.
система РА88, которая одновременно предсказывает вероятность более чем 100 фармакологических эффектов и механизмов действия вещества на основе его структурной формулы. Эффективность применения этого подхода к планированию скрининга составляет около 800%, а точность компьютерного прогноза на 300% превосходит предсказание экспертов.
Итак, одним из конструктивных инструментов получения новых знаний и решений в медицине является метод математического моделирования. Процесс математизации медицины – частое проявление взаимопроникновения научных знаний, повышающее эффективность лечебно-профилактической работы.
4. Математическая модель «хищники-жертвы»
Впервые в биологии математическую модель периодического изменения числа антагонистических видов животных предложил итальянский математик В. Вольтерра с сотрудниками. Модель, предложенная Вольтерра, явилась развитием идеи, намеченной в 1924 году А. Лоттки в книге "Элементы физической биологии". Поэтому эта классическая математическая модель известна как модель "Лоттки-Вольтерра".
Хотя в природе отношения антагонистических видов более сложные, чем в модели, тем не менее они являются хорошей учебной моделью, на которой можно изучать основные идеи математического моделирования.
Итак, задача : в некотором экологически замкнутом районе живут два вида животных (например, рыси и зайцы). Зайцы (жертвы) питаются растительной пищей, имеющейся всегда в достаточном количестве (в рамках данной модели не учитывается ограниченность ресурсов растительной пищи). Рыси (хищники) могут питаться только зайцами. Необходимо определить, как будет меняться численность жертв и хищников с течением времени в такой экологической системе. Если популяция жертв увеличивается, вероятность встреч хищников с жертвами возрастает, и, соответственно, после некоторой временной задержки, растет популяция хищников. Эта достаточно простая модель вполне адекватно описывает взаимодействие между реальными популяциями хищников и жертв в природе.
Теперь приступим к составлению дифференциальных уравнений. Обо-
значим число жертв через N, а число хищников через M. Числа N и M являются функциями времени t . В нашей модели учтем следующие факторы:
а) естественное размножение жертв; б) естественная гибель жертв;
в) уничтожение жертв за счет поедания их хищниками; г) естественное вымирание хищников;
д) увеличение числа хищников за счет размножения при наличии пищи.
Так как речь идет о математической модели, то задачей является получение уравнений, в которые входили бы все намеченные факторы и которые описывали бы динамику, то есть изменение числа хищников и жертв со временем.
Пусть за некоторое время t количество жертв и хищников изменится на ∆N и ∆M. Изменение числа жертв ∆N за время ∆t определяется, во-первых, увеличением в результате естественного размножения (которое пропорционально имеющемуся количеству жертв):
где В – коэффициент пропорциональности, характеризующий скорость естественного вымирания жертв.
В основе вывода уравнения, описывающего уменьшение числа жертв изза поедания их хищниками, лежит идея о том, что чем чаще происходит их встреча, тем быстрее уменьшается число жертв. Ясно также, что частота встреч хищников с жертвой пропорционально и числу жертв и числу хищников, то
Поделив левую и правую часть уравнения (4) на ∆t и перейдя к пределу при ∆t→0 , получим дифференциальное уравнение первого порядка:
Для того, чтобы решить это уравнение, нужно знать, как меняется число хищников (М ) со временем. Изменение числа хищников (∆М ) определяется увеличением из-за естественного размножения при наличии достаточного количества пищи (М 1 = Q∙N∙M∙∆t ) и уменьшением из-за естественного вымирания хищников (M 2 = - P∙M∙∆t ):
M = Q∙N∙M∙∆t - P∙M∙∆t |
Из уравнения (6) можно получить дифференциальное уравнение:
Дифференциальные уравнения (5) и (7) представляют собой математическую модель "хищники-жертвы". Достаточно определить значения коэффици-

ентов A, B, C, Q, P и математическую модель можно использовать для решения поставленной задачи.
Проверка и корректировка математической модели. В данной лабора-
торной работе предлагается кроме просчета наиболее полной математической модели (уравнения 5 и 7), исследовать более простые, в которых что-либо не учитывается.
Рассмотрев пять уровней сложности математической модели, можно "почувствовать" этап проверки и корректировки модели.
1-ый уровень – в модели учтено для "жертв" только их естественное размножение, "хищники" отсутствуют;
2-ой уровень – в модели учтено для "жертв" их естественное вымирание, "хищники" отсутствуют;
3-ий уровень – в модели учтены для "жертв" их естественное размножение
и вымирание, "хищники" отсутствуют;
4-ый уровень – в модели учтены для "жертв" их естественное размножение
и вымирание, а также поедание "хищниками", но число "хищников" остается неизменным;
5-ый уровень – в модели учтены все обсуждаемые факторы.
Итак, имеем следующую систему дифференциальных уравнений:
где М – число "хищников"; N – число "жертв";
t – текущее время;
A – скорость размножения "жертв"; C – частота встреч "хищники-жертвы"; B – скорость вымирания "жертв";
Q – размножение "хищников";
P – вымирание "хищников".
1-ый уровень: М = 0, В = 0; 2-ой уровень: М = 0, А = 0; 3-ий уровень: М = 0; 4-ый уровень: Q = 0, Р = 0;
5-ый уровень: полная система уравнений.
Подставляя значения коэффициентов в каждый уровень, будем получать разные решения, например:
Для 3-его уровня значение коэффициента М=0 , тогда

решая уравнение получим
Аналогично для 1-го и 2-го уровней. Что касается 4-го и 5-го уровней, то здесь необходимо решать систему уравнений методом Рунге-Кутта. В результате получим решение математических моделей данных уровней.
II. РАБОТА СТУДЕНТОВ ВО ВРЕМЯ ПРАКТИЧЕСКОГО ЗАНЯТИЯ
Задание 1 . Устно-речевой контроль и коррекция усвоения теоретического материала занятия. Сдача допуска к занятию.
Задание 2 . Выполнение лабораторной работы, обсуждение полученных результатов, оформление конспекта.
Выполнение работы
1. С рабочего стола компьютера вызвать программу "Лаб. №6", щелкнув по соответствующему ярлыку два раза левой клавишей "мыши".
2. Щелкнуть дважды левой клавишей "мыши" по ярлыку "PREDATOR".
3. Выбрать ярлык "PRED" и повторить вызов программы левой клавишей "мыши" (щелкнув дважды).
4. После титульной заставки нажать "ENTER".
5. Моделирование начинать с 1-го уровня.
6. Ввести год, с которого будет проводиться анализ модели: например, 2000
7. Выбрать временные интервалы, например, в течение 40 лет, через 1 год (затем через 4 года).
2-ой уровень: B = 0.05; N0 = 200;
3-ий уровень: A = 0.02; B = 0.05; N = 200;
4-ый уровень: A = 0.01; B = 0.002; C = 0.01; N0 = 200; M = 40; 5-ый уровень: A = 1; B = 0.5; C = 0.02; Q = 0.002; P = 0.3; N0 = 200;
9. Подготовить письменный отчет по работе, который должен содержать уравнения, графики, результаты расчета характеристик модели, выводы по проделанный работе.
Задание 3. Контроль конечного уровня знаний:
а) устно-речевой отчет за выполненную лабораторную работу; б) решение ситуационных задач; в) компьютерное тестирование.
Задание 4. Задание на следующее занятие: раздел и тема занятия, согласование тем реферативных докладов (объем доклада 2-3 стр., регламент 5-7 мин.).
Здесь в отличие от (3.2.1) знаки (-012) и (+a2i) разные. Как и в случае конкуренции (система уравнений (2.2.1)), начало координат (1) для этой системы является особой точкой типа «неустойчивый узел». Три других возможных стационарных состояния:

Биологический смысл требует положительности величин Х у х 2. Для выражения (3.3.4) это означает, что
В случае, если коэффициент внутривидовой конкуренции хищников а ,22 = 0, условие (3.3.5) приводит к условию ai2
Возможные типы фазовых портретов для системы уравнений (3.3.1) представлены на рис. 3.2 a-в. Изоклины горизонтальных касательных представляют собой прямые
а изоклины вертикальных касательных - прямые
Из рис. 3.2 видно следующее. Система хищник -жертва (3.3.1) может иметь устойчивое положение равновесия, в котором популяция жертв полностью вымерла (х = 0) и остались только хищники (точка 2 на рис. 3.26). Очевидно, такая ситуация может реализоваться лишь в случае, если кроме рассматриваемого вида жертв х хищник Х 2 имеет дополнительные источники питания. Этот факт в модели отражается положительным членом в правой части уравнения для хз. Особые точки (1) и (3) (рис. 3.26) являются неустойчивыми. Вторая возможность - устойчивое стационарное состояние, в котором популяция хищников полностью вымерла и остались одни жертвы - устойчивая точка (3) (рис. 3.2а). Здесь особая точка (1) - также неустойчивый узел.
Наконец, третья возможность - устойчивое сосуществование популяций хищника и жертвы (рис. 3.2 в), стационарные численности которых выражаются формулами (3.3.4). Рассмотрим этот случай подробнее.
Предположим равенство нулю коэффициентов внутривидовой конкуренции (аи = 0, i = 1, 2). Предположим также, что хищники питаются только жертвами вида х и в отсутствие их вымирают со скоростью С2 (в (3.3.5) С2
Проведем подробное исследование этой модели, воспользовавшись обозначениями, наиболее широко принятыми в литературе. Переобо-

Рис. 3.2. Расположение главных изоклин на фазовом портрете вольтерров- ской системы хищник жертва при разном соотношении параметров: а - о» б -
С I С2 С2
1, 3 - неустойчивые, 2 - устойчивая особая точка; в -
1, 2, 3 - неустойчивые, 4 - устойчивая особая точка значим
Система хищник-жертва в этих обозначениях имеет вид:

Свойства решений системы (3.3.6) будем исследовать на фазовой плоскости N
1
ON
2
Система имеет два стационарных решения. Их легко определить, приравняв нулю правые части системы. Получим:

Отсюда стационарные решения:

Рассмотрим подробнее второе решение. Найдем первый интеграл системы (3.3.6), не содержащий t. Умножим первое уравнение на -72, второе - на -71 и результаты сложим. Получим:
Теперь разделим первое уравнение на N и умножим на € 2, а второе разделим на JV 2 и умножим на е. Результаты снова сложим:
Сравнивая (3.3.7) и (3.3.8), будем иметь:

Интегрируя, получим:

Это и есть искомый первый интеграл. Таким образом, система (3.3.6) является консервативной, поскольку имеет первый интеграл движения, величину, представляющую собой функцию переменных системы N и N 2 и не зависящую от времени. Это свойство позволяет конструировать для вольтерровских систем систему понятий, аналогичную статистической механике (см. гл. 5), где существенную роль играет величина энергии системы, неизменная во времени.
При каждом фиксированном с > 0 (что соответствует определенным начальным данным) интегралу соответствует определенная траектория на плоскости N 1 ON 2 , служащая траекторией системы (3.3.6).
Рассмотрим графический способ построения траектории, предложенный самим Вольтерра. Заметим, что правая часть формулы (3.3.9) зависит только от Д г 2, а левая - только от N. Обозначим
Из (3.3.9) следует, что между X и Y имеется пропорциональная зависимость
На рис. 3.3 изображены первые квадранты четырех систем координат XOY, NOY , N 2 OX и Д Г 10N 2 так, чтобы все они имели общее начало координат.
В левом верхнем углу (квадрант NOY) построен график функции (3.3.8), в правом нижнем (квадрант N 2 OX) - график функции Y. Первая функция имеет min при Ni = а вторая - max при N 2 = ?-
Наконец, в квадранте XOY построим прямую (3.3.12) для некоторого фиксированного С.
Отметим точку N на оси ON . Этой точке соответствует определенное значение Y(N 1), которое легко найти, проведя перпендикуляр

Рис. 3.3.
через N до пересечения с кривой (3.3.10) (см. рис. 3.3). В свою очередь, значению К(Д^) соответствует некоторая точка М на прямой Y = сХ и, следовательно, некоторое значение X(N) = Y(N)/c, которое можно найти, проведя перпендикуляры AM и MD. Найденному значению (эта точка отмечена на рисунке буквой D) соответствуют две точки Р и G на кривой (3.3.11). По этим точкам, проводя перпендикуляры, найдем сразу две точки Е" и Е ", лежащие на кривой (3.3.9). Их координаты:

Проводя перпендикуляр AM , мы пересекли кривую (3.3.10) еще в одной точке В. Этой точке соответствуют те же Р и Q на кривой (3.3.11) и те же N и Щ. Координату N этой точки можно найти, опустив перпендикуляр из В на ось ON. Таким образом, мы получим точки F" и F", также лежащие на кривой (3.3.9).
Исходя из другой точки N, тем же самым образом получим новую четверку точек, лежащих на кривой (3.3.9). Исключение составит точка Ni = ?2/72- Исходя из нее, получим только две точки: К и L. Это будут нижняя и верхняя точки кривой (3.3.9).
Можно исходить не из значений N , а из значений N 2 . Направляясь от N 2 к кривой (3.3.11), поднимаясь затем до прямой У = сХ, а оттуда пересекая кривую (3.3.10), также найдем четыре точки кривой (3.3.9). Исключение составит точка No = ?1/71- Исходя из нее, получим только две точки: G и К. Это будут самая левая и самая правая точки кривой (3.3.9). Задавая разные N и N 2 и получив достаточно много точек, соединив их, приближенно построим кривую (3.3.9).
Из построения видно, что эго замкнутая кривая, содержащая внутри себя точку 12 = (?2/721 ?1/71)» исходящая из определенных начальных данных N ю и N20. Взяв другое значение С, т.е. другие начальные данные, получим другую замкнутую кривую, не пересекающую первую и также содержащую точку (?2/721 ?1/71)1 внутри себя. Таким образом, семейство траекторий (3.3.9) есть семейство замкнутых линий, окружающих точку 12 (см. рис. 3.3). Исследуем тип устойчивости этой особой точки, воспользовавшись методом Ляпунова.
Так как все параметры е 1, ?2, 71,72 положительны, точка (N[расположена в положительном квадранте фазовой плоскости. Линеаризация системы вблизи этой точки дает:

Здесь n(t) и 7i2(N1, N 2 :
Характеристическое уравнение системы (3.3.13):

Корни этого уравнения чисто мнимые:

Таким образом, исследование системы показывает, что траектории вблизи особой точки представлены концентрическими эллипсами, а сама особая точка - центр (рис. 3.4). Рассматриваемая модель Вольтерра и вдали от особой точки имеет замкнутые траектории, хотя форма этих траекторий уже отличается от эллипсоидальной. Поведение переменных Ni, N 2 во времени показано на рис. 3.5.

Рис. 3.4.

Рис. 3.5. Зависимость численности жертвы N i и хищника N 2 от времени
Особая точка типа центр устойчива, но не асимптотически. Покажем на данном примере, в чем это заключается. Пусть колебания Ni(t) и ЛГгМ происходят таким образом, что изображающая точка движется по фазовой плоскости по траектории 1 (см. рис. 3.4). В момент, когда точка находится в положении М, в систему извне добавляется некоторое количество особей N 2, такое, что изображающая точка переходит скачком из точки М в точку Л/". После этого, если система снова предоставлена самой себе, колебания Ni и N 2 уже будут происходить с большими амплитудами, чем прежде, и изображающая точка двигается по траектории 2. Это и означает, что колебания в системе неустойчивы: они навсегда изменяют свои характеристики при внешнем воздействии. В дальнейшем мы рассмотрим модели, описывающие устойчивые колебательные режимы, и покажем, что на фазовой плоскости такие асимптотические устойчивые периодические движения изображаются при помощи предельных циклов.
На рис. 3.6 изображены экспериментальные кривые - колебания численности пушных зверей в Канаде (по данным компании Гудзонова залива). Эти кривые построены на основании данных по числу заготовленных шкурок. Периоды колебаний численности зайцев (жертв) и рысей (хищников) примерно одинаковы и порядка 9 10 лет. При этом максимум численности зайцев опережает, как правило, максимум численности рысей на один год.
Форма этих экспериментальных кривых значительно менее правильная, чем теоретических. Однако в данном случае достаточно того, что модель обеспечивает совпадение наиболее существенных характеристик теоретических и экспериментальных кривых, г.е. величин амплитуды и сдвига фаз между колебаниями численностей хищников и жертв. Гораздо более серьезным недостатком модели Вольтерра является неустойчивость решений системы уравнений. Действительно, как уже говорилось выше, любое случайное изменение численности того или другого вида должно привести, следуя модели, к изменению амплитуды колебаний обоих видов. Естественно, что в природных условиях животные подвергаются бесчисленному количеству таких случайных воздействий. Как видно из экспериментальных кривых, амплитуда колебаний численностей видов мало изменяется от года к году.
Модель Вольтерра - эталонная (базовая) для математической экологии в той же мере, в какой модель гармонического осциллятора является базовой для классической и квантовой механики. При помощи этой модели на основе очень упрощенных представлений о характере закономерностей, описывающих поведение системы, сугубо математи-
Глава 3

Рис. 3.6. Кинетические кривые численности пушных зверей по данным пуш ной компании Гудзонова залива (Сетон-Томсон, 1987) ческими средствами было выведено заключение о качественном характере поведения такой системы - о наличии в такой системе колебаний численности популяции. Без построения математической модели и ее использования такой вывод был бы невозможен.
В рассмотренном нами выше самом простом виде системе Воль- терра присущи два принципиальных и взаимосвязанных недостатка. Их «устранению» посвящена обширная эколого-математическая литература. Во-первых, включение в модель любых, сколь угодно малых, дополнительных факторов качественным образом меняет поведение системы. Второй «биологический» недостаток модели заключается в том, что в нее не включены принципиальные свойства, присущие любой паре взаимодействующих по принципу хищник-жертва популяций: эффект насыщения хищника, ограниченность ресурсов хищника и жертвы даже при избытке жертвы, возможность минимальной численности жертв, доступных для хищника, и пр.
С целью устранения этих недостатков были предложены разными авторами различные модификации системы Вольтерра. Наиболее ин- тересные из них будут рассмотрены в разделе 3.5. Здесь остановимся лишь на модели, учитывающей самоограничения в росте обеих популяций. На примере этой модели наглядно видно, как может меняться характер решений при изменении параметров системы.
Итак, рассматривается система

Система (3.3.15) отличается от ранее рассмотренной системы (3.3.6) наличием в правых частях уравнений членов вида -7uNf,
Эти члены отражают тот факт, что численность популяции жертв не может расти до бесконечности даже в отсутствие хищников в силу ограниченности пищевых ресурсов, ограниченности ареала существования. Такие же «самоограничения» накладываются и на популяцию хищников.
Для нахождения стационарных численностей видов iVi и N 2 приравняем к нулю правые части уравнений системы (3.3.15). Решения с нулевыми значениями численностей хищников или жертв не будут нас сейчас интересовать. Поэтому рассмотрим систему алгебраических
уравнений
 Ее решение
Ее решение
дает нам координаты особой точки. На параметры системы здесь следует положить условие положительности стационарных численностей: N > 0 и N 2 > 0. Корни характеристического уравнения системы, линеаризованной в окрестности особой точки (3.3.16):

Из выражения для характеристических чисел видно, что если выполнено условие
то численности хищников и жертв совершают во времени затухающие колебания, система имеет ненулевую особую точку устойчивый фокус. Фазовый портрет такой системы изображен на рис. 3.7 а.
Допустим, что параметры в неравенстве (3.3.17) так изменяют свои значения, что условие (3.3.17) обращается в равенство. Тогда характеристические числа системы (3.3.15) равны, а ее особая точка будет лежать на границе между областями устойчивых фокусов и узлов. При изменении знака неравенства (3.3.17) на обратный особая точка становится устойчивым узлом. Фазовый портрет системы для этот случая представлен на рис. 3.76.
Как и в случае одной популяции, для модели (3.3.6) можно разработать стохастическую модель, но для нее нельзя получить решение в явном виде. Поэтому мы ограничимся общими рассуждениями. Допустим, например, что точка равновесия находится на некотором расстоянии от каждой из осей. Тогда для фазовых траекторий, на которых значения JVj, N 2 остаются достаточно большими, вполне удовлетворительной будет детерминистическая модель. Но если в некоторой точке


Рис. 3.7. Фазовый портрет системы (3.3.15): а - при выполнении соотношения (3.3.17) между параметрами; б - при выполнении обратного соотношения между параметрами
фазовой траектории какая-либо переменная не очень велика, то существенное значение могут приобрести случайные флуктуации. Они приводят к тому, что изображающая точка переместится на одну из осей, что означает вымирание соответствующего вида. Таким образом, стохастическая модель оказывается неустойчивой, так как стохастический «дрейф» рано или поздно приводит к вымиранию одного из видов. В такого рода модели хищник в конечном счете вымирает, это может произойти либо случайно, либо вследствие того, что сначала элиминируется популяция его жертвы. Стохастическая модель системы хищник- жертва хорошо объясняет эксперименты Гаузе (Гаузе, 1934; 2000), в которых инфузория Paramettum candatum служила жертвой для другой инфузории Didinium nasatum - хищника. Ожидавшиеся согласно детерминистическим уравнениям (3.3.6) равновесные численности в этих экспериментах составляли примерно всего но пять особей каждого вида, так что нет ничего удивительного в том, что в каждом повторном эксперименте довольно быстро вымирали либо хищники, либо жертвы (а за ними и хищники).
Итак, анализ вольтерровских моделей взаимодействия видов показывает, что, несмотря на большое разнообразие типов поведения таких систем, незатухающих колебаний численности в модели конкурирующих видов не может быть вовсе. В модели хищник жертва незатухающие колебания появляются вследствие выбора специальной формы уравнений модели (3.3.6). При этом модель становится негрубой, что свидетельствует об отсутствии в такой системе механизмов, стремящихся сохранить ее состояние. Однако в природе и эксперименте такие колебания наблюдаются. Необходимость их теоретического объяснения послужила одной из причин для формулировки модельных описаний в более общем виде. Рассмотрению таких обобщенных моделей посвящен раздел 3.5.



